Concerns over the safety of vaccines and effectiveness of non-pharmaceutical control measures have led to and will lead to worse outcomes for COVID, especially in the US, where such resistance has led to tragic devastation. Therefore, any over interpretation of worst case scenarios will lead to self-fulfilling apocalyptic outcomes. This can only be prevented by open and honest assessment of risk/cost/benefits of any such approaches https://www.medrxiv.org/content/10.1101/2021.01.29.21250653v1.full.pdf. The latest most concerning challenges to vaccines are their potential to instigate autoimmunity, allergic reactivity, their ability to enhance infectivity, and their loss of effectiveness because of the appearance and rapid spread of mutant strains of SARS-CoV-2. Another more common place and older concern for attenuated vaccines is their possible reversion to pathogenic forms. I will start with this last concern first because it has some well-documented clinical examples. The earliest was with the first vaccine, smallpox vaccine, cowpox or now vaccinia (DNA from an early 1902 sample of smallpox vaccine showed that it was 99.7% similar to horsepox virus, now extinct). This vaccine is a close attenuated relative of smallpox virus, usually only causing localized lesions, skin vesicles, followed by erosions and ulceration, at most resulting in a single scar. However, it can cause generalized vaccinia, with widespread reddening, inflammation and pealing of the skin and on very rare occasion heart damage and even death (eczema vaccinatum, accidental inoculation, and progressive vaccinia). The spread of the virus from inoculation sites during active infections to others, especially amongst children, was common in the past. But we must never forget that without this vaccine, smallpox could have never been eradicated. We would still be living with its up to 30% mortality and life-long disfigurement.
A lesser known reversion was seen with Venezuelan Equine Encephalitis modified live virus vaccine, TC-83 virus, was developed as an anti-biological warfare vaccine in the early 1960’s at USAMRIID (US Army Medical Research Institute of Infectious Diseases) and transitioned to a commercial horse vaccine in the 1970’s and eventually was replaced with a killed vaccine. VEEV was attenuated into TC-83 by 83 serial passages in guinea pig heart cells. It was first tested extensively in horses during the 1971 Texas VEE epizootic. Although the vaccine produces viremia, fever and leucopenia in horses, neutralizing antibodies are generated with protection from VEEV challenge. TC-83 produces seroconversion in about 80% of humans (neutralizing antibody titers ≥20) but mild to moderate flu-like symptoms. TC-83 is the result of only 2 attenuating mutations among the 12 mutations that accompanied in vitro passage of the Trinidad donkey strain. These mutations could revert in vaccinees, which also presented a risk of epizootic amplification. To produce a more stably attenuated VEEV vaccine candidate, the virulent Trinidad donkey genome was cloned in cDNA and attenuated by inserting either point mutations or a PE2 cleavage-signal mutation combined with an E1 gene resuscitating mutation. The latter strain, called V3526, is safe and immunogenic for mice and nonhuman primates, and appears superior to TC-83 in rodents. V3526 has a lower risk for environmental transmission and distribution. TC-83 was transmitted by mosquito vectors and was isolated in mosquitoes collected in Louisiana during the 1971 VEE epizootic/epidemic. Efforts were made to improve its safety by genetic modification to prevent its ability to infect insects. This recombinant virus further attenuated TC-83, but was completely unable to infect mosquito cells or mosquitoes in vivo. Currently, commercial equine vaccines marketed in the U.S. consist of inactivated TC-83, often combined with inactivated eastern and western equine encephalitis viruses.
“The small and simple VEEV has one more trick which it shares with the plant Tobacco Mosaic Virus; it can resurrect itself from broken pieces. The nsP4 sequence in VEEV is very similar to the replicases of three plant viruses, including Tobacco Mosaic Virus. The Brooks Lab performed a critical experiment using the RetroXpress (TM) system of Clontech derived from a retrovirus which could transfer genes but could not infect and replicate, my group at Brooks constructed a new virus that contained the nsP4 sequence inserted into this platform and expected that the new virus could infect cells but not replicate. It not only infected (transfected) marker genes into the target mammalian cells but also formed plaques of transformed cells (those which resembled cancer cells and formed uninhibited growth mounds of cells). These grew and multiplied in the cell culture. A new complete infectious virus had formed. How? Tobacco Mosaic Virus has been shown to repair and reactivate after exposure to UV with the help of an unknown plant nucleic acid repair system (Multiplicity Reactivation: Recombination). Since we inserted only the nsP4 protein into the host cells co-infected with the defective retrovirus, we proposed that the new virus used cellular RNA dependent RNA polymerase in conjunction with the expressed nsP4 to form a repair complex which constructed a chimera virus from the retroviral pieces and other unknown viral nucleic acid components inherently present. Perhaps, the infections by killed virus from killed vaccines previously reported were not just because of contamination with whole live virus but with components which survived the formalin damage and were reassembled into whole restored infectious virus, albeit at a less effective infectious dose of original broken pieces. Intact VEEV will produce a transmissible infection with as little as 12 PFU (plaque forming units equivalent to 10 virions for a multiplicity of infection of 1 but more likely 10X10 = 100 virions). I have speculated that these broken viruses, if reconstituted from broken virions or free RNA would require 10e6 units to make one intact virion.” — The Black Dragon Trilogy by JOHNATHAN KIEL. https://a.co/gtIO9aR
The third example of a problematic attenuated live vaccine still causing trouble is Sabin modified live polio vaccine. The first, vaccine-associated paralytic poliomyelitis (VAPP) was recognized in the early 1960s. VAPP is a very rare and occurs sporadically when a dose of oral live polio vaccine reverts to neurovirulence and causes paralysis in the vaccine recipient or a nonimmune contact. VAPP can be in immunologically normal vaccine recipients and their contacts as well as among persons with immunodeficiencies, defects in antibody production. There were live vaccine-associated outbreaks in Madagascar and South Sudan in 2014. Eight newly identified persons in six countries were found to excrete vaccine virus, and a patient in the United Kingdom was still excreting the virus in 2014 after more than 28 years. Reported by ProMed on 15 Dec 2020 were three new polio cases in Sierra Leone for the first time in 10 years. The cases were recorded in Kambia, Tonkolili, and Western Area Rural District. The cases were not from the wild poliovirus strain, but were vaccine derived. Only 2 countries worldwide continue to have wild poliovirus transmission: Pakistan and Afghanistan; thanks to worldwide polio vaccination. Because of violent resistance to polio vaccination in these two countries, polio persists and cannot be eradicated. Killed Salk polio vaccination is now once again being used with no chance of reversion because it is a killed vaccine, as the risk of live Sabin vaccine now out weighs its benefits.
The severe reactions to vaccines (anaphylaxis) are extremely rare for humans and animals and are expected to be no worse for COVID vaccines. They are often, when they do occur associated with adjuvants and additives, rather than the primary immunogen, except for perhaps some vaccines in animals, like that for leptospirosis (actually a bacterin).
Autoimmunity is related to allergic reaction but distinct, especially in regards to SARS-CoV-2 pathology. I have discussed the immune response to the virus in detail in previous posts. As a reminder, the virus has the unusual association of autoantibody against interferons as a marker of severe disease. Also, unlike other animal coronaviruses, SARS-CoV-2 is more severe in adults than children, in general. The animal ones are more severe in the young than old. What is a plausible explanation, or hypothesis? Children have a more robust innate immune response, including interferon, than an adaptive immune (antibody) response than adults, but they have higher amounts of circulating soluble interleukin 17 receptors which help reduce the cellular innate pro-inflammatory response mediated in immune cells through membrane-bound IL-17 receptors. I have pointed out, in an earlier post, that the uncontrollable innate immune response yields reactive oxygen species that cause considerable local collateral host cell damage and death. Therefore, taking out interferon, and the subsequent wide distribution of infectious virus, then leads to antibody against virally infected cells which directs antibody-dependent cell cytotoxicity, causing a whole host inflammatory response, and collateral cellular, tissue to whole organ damage and failure. The pro-inflammatory activity of whole virus leads to a by-stander adjuvant effect which can activate forbidden B cells to produce the self antibody, i.e. anti-interferon. In adult immune systems, having seen more related viruses than the naive immune systems of children, this is more likely to happen. A good humoral and cellular immune response, triggered by a vaccine directed against specific antigens that direct neutralizing antibody response avoids this outcome because it takes out virus before it can spread widely in the body.
This line of thinking leads to a more recent disconcerting response to the virus that involves a minor antibody directed to a critical part of the binding portion of the viral spike protein (S1) https://www.biorxiv.org/content/10.1101/2020.12.18.423358v1?fbclid=IwAR3D1MJ-SyW9wLeqrRV6pdSP6ToUwpaLjbB5LK2oA4B0b9iYjJ4RTvYs_PQ. The paper provides evidence that minor antibodies from COVID patients’ serum bind to part of the S1 spike protein that enhances the binding to ACE2 viral receptors and uptake of infectious virus in cell culture in established cell lines. They further raise issue that a vaccine could enhance viral uptake and disease by extension of this observation to the whole human. Unfortunately, such a stretch will provide support to those who oppose vaccination. This leap may be moot from the get go because patients have circulating soluble ACE2, derived from proteolytically cleaved membrane bound ACE2. First, there is no evidence that any vaccine enhances viral infectivity in vivo. Second, these soluble receptors would first see and soak up systemic virus, especially if its binding were enhanced by the antibody. Then, immune cells would take up these immune complexes and dispose of the virus. In addition, the antigens selected for the vaccines are specifically selected to optimize induction of neutralizing antibody.
The vaccines released for use so far have had exceptional efficacy in generating neutralizing antibody. The fear of a mutation in SARS-CoV-2 has come to fruition but those so far such as in mink, have increased efficacy of transmission and infection but not pathogenicity. Coronaviruses have a large RNA genome and can therefore undergo more mutations without significant functional change than smaller genome RNA viruses. They also have a proof reading polymerase to reduce replication mistakes (mutations) unlike other error prone RNA viruses like HIV. However, they can undergo selection pressure for advantageous mutants, when necessary, generated by mutations mediated by Nsp14 exonuclease in independent coronaviruses, resulting in a 15–21-fold decrease in replication fidelity. Therefore, the presence of this exonuclease can clearly influence viral RNA mutation rates and the generation of quasi species when needed. Therefore, there is a continuous tension balance between the polymerase and the exonuclease to only produce mutations that are favored by environmental selection pressure. The latest of greatest concern, VOC 202012/01 variant, occurred in the UK. This variant has a mutation in the receptor binding domain (RBD) of the spike protein at position 501, where the amino acid asparagine (N) has been replaced with tyrosine (Y). The shorthand of the mutation is N501Y, also designated as S:N501Y, because it is in the spike protein. This variant carries many other mutations, 14 amino acid (AA) mutations, 6 non-AA altering mutations, and 3 deletions, notably including 69/70 deletion: this double deletion has occurred spontaneously many times, and likely leads to a change in the shape of the spike protein; P681H: near the S1/S2 furin cleavage site, a site with high variability in coronaviruses, which has also emerged spontaneously multiple times; ORF8 stop codon (Q27stop), a mutation not in the spike protein but in a different gene (in open reading frame 8), the function of which is unknown. Similar mutations have occurred in the past such as in Singapore, in which one strain with this type of mutation emerged and disappeared. Preliminary analysis in the UK indicates this variant is significantly more transmissible than previously circulating variants, with an estimated potential to increase the reproductive number (R) by 0.4 or greater with an estimated increased transmissibility of up to 70%. There is no indication, at this point, of increased disease severity associated with the new variant. A few cases of this new variant have to date been reported in Denmark and the Netherlands. Another similar mutant has arisen in South Africa, with structural changes in the spike protein, which is still being evaluated for impact. However, in a recently released study, pending peer review, the Pfizer vaccine has been shown to still induce neutralizing protection to the mutants https://www.biorxiv.org/content/10.1101/2021.01.07.425740v1.full.pdf. One must remember that mutations in the spike protein are not limitless. They must conserve essential structure that allows binding and entry of the virus https://science.sciencemag.org/content/sci/371/6526/284.full.pdf. The ACE2 viral receptors do not mutate as fast as the virus so they limit what variations in the virus are still tolerable for infection. If coronavirus behavior is to hold true, the most efficiently transmitted virus with the lowest mortality necessary to eliminate less than ideal virus replicating hosts, and that which does not preferentially infect much more than 10% of the susceptible population, especially when simple non-pharmaceutical methods as distancing, good ventilation and wearing of masks, are so effective, will prevail to maintain itself in the general population indefinitely. This persistence at a low percentage of the susceptible population is the nature of coronaviruses in animals.
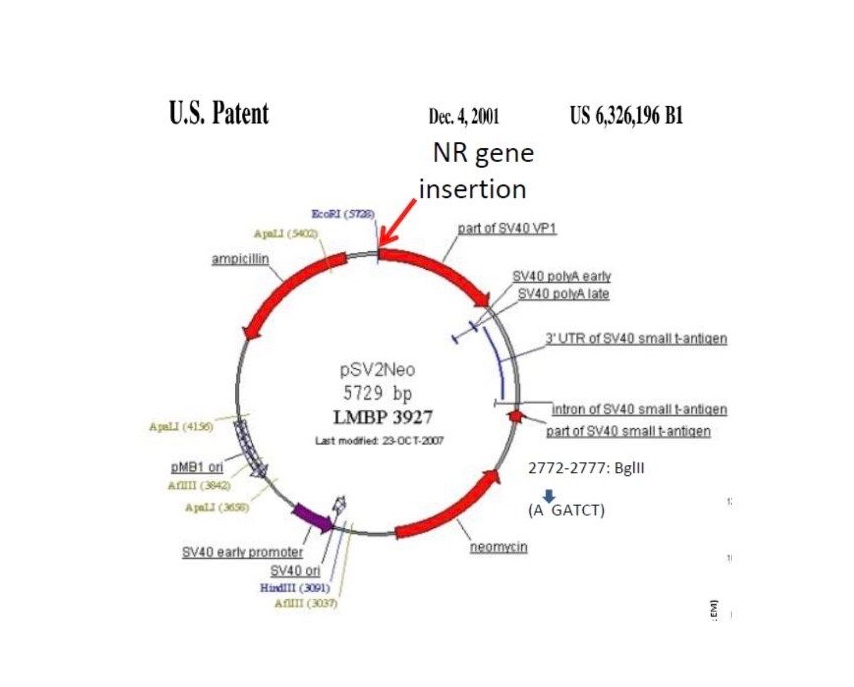
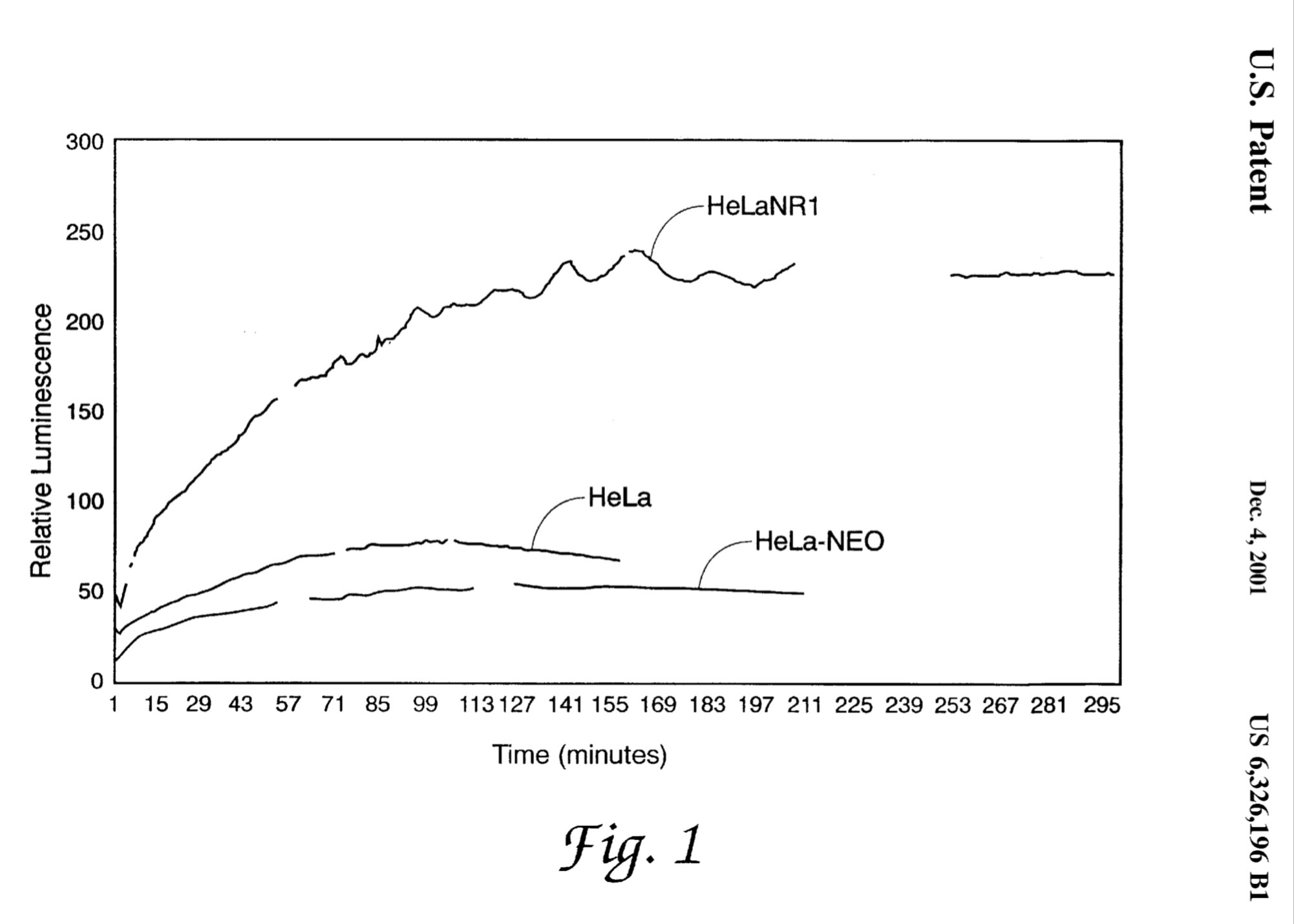
One last note, the Brooks USAF AFRL CPT groups’ seeking of aptamer-based anti-pathogens to bind toxic components, inhibit target cell binding and uptake, and re-direct existing immunity, natural or vaccine induced, rapidly toward new, emerging and re-emerging pathogens, including viruses, was designed to address outbreaks like COVID without always having to resort to the Herculean efforts that have been necessary to develop COVID vaccines. The failure or inability to apply the same efforts, as we have had to to rapidly develop COVID vaccines, to the insidious smoldering problem of antimicrobial (antibiotic) resistance was also the motivation for developing this new technology, as well as to relieve the costs and socioeconomic impacts of addressing these problems by conventional, albeit modern technological, versions of biomedical approaches.