As stated in an earlier post, it is much more difficult to mutate past many different antiviral antibodies or different simultaneous multiple drug therapies. For example, in 1995, a combination drug treatment known as the “AIDS cocktail” was introduced which made AIDS a survivable, chronic disease. This type of therapy is known as highly active antiretroviral therapy (HAART), also called combination antiretroviral therapy (cART). HIV is a virus much more prone to mutations than SARS-CoV-2. Our own research, demonstrated that a mutagen (2-chloroethylethylsulfide) could make bacteria resistant to one but not two antibiotics at the same time in the same microbes without substantial mutations that largely proved lethal for the bacteria (ampicillin and 3-amino-L-tyrosine: Patent #: 5,902,728; date: May11,1999). As the number of mutations increased in the microbial population in order to achieve both ampicillin resistance and 3-amino-L-tyrosine/nitrate resistance, the bacteria became less robust and sacrificed their competitiveness to less resistant subpopulations. The JM109/pIC20RNR1.1 E. coli, genetically modified bacteria by a plasmid, was designed to have conflicting selection pressures when grown in 3-AT medium containing both ampicillin and 3-amino-L-tyrosine/nitrate. The biosynthesis of the DALM polymer was driven by the ampicillin resistance expressed from the same plasmid which contained the gene for enhanced DALM biosynthesis. However, driving DALM synthesis led to lethality. Loss of the ampicillin resistance would spare the bacteria from DALM lethality, but lead to their death by the ampicillin. Therefore, there needed to be a set of simultaneous mutations to spare the bacteria from both these diametrically opposed lethal effects. A similar response would prevent SARS-CoV-2 from escape mutations if hit by several different antiviral approaches simultaneously. The number of mutations in SARS-CoV-2 increase by chance as the circulating population of viruses increases and they become available for natural selection for traits that increase their survivability and transmission. Conversely, limiting their spread and numbers by both pharmaceutical (vaccines and antivirals) and non-pharmaceutical means (masks and distancing) greatly reduce these possibilities. Finally, I illustrate below how the appearance of variants, which evade vaccines and treatments, can be countered by aptamer adapters that restore the effectiveness of these vaccines and treatments.

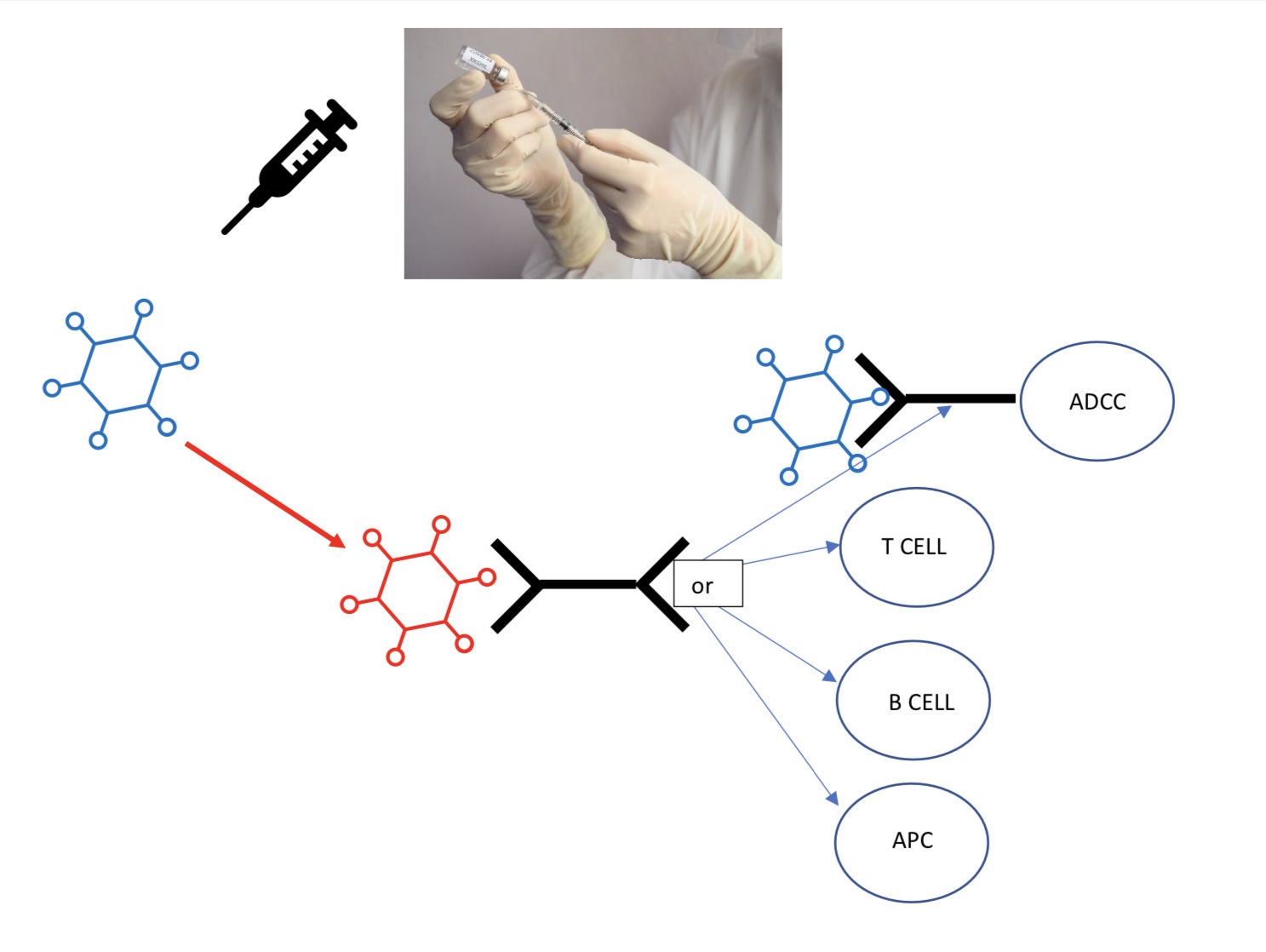
Re-Directing an old vaccine against a new variant of a virus. The blue virus was vaccinated against, then the circulating virus mutated into the red virus which is not protected against by the previous vaccine. The red virus is isolated, then nucleic acid aptamers are selected in vitro against the new virus. Another set of aptamers are selected against ligands on existing antibody against the original blue virus, or against ligands on cells in the immune system to recruit them to attack the virus (NK=Natural Killer cells, cytotoxic T cells, helper T cells and associated B cells). These aptamer sequences can be fused together to provide an adapter (>~~<) which links the binding of the variant virus to mediator (APC, Antigen Presenting Cells; dendrocytes) and effector cells in the immune system. The linkage to pre-existing antibody to the predecessor virus (>~~) allows neutralization of the variant virus and recruitment of Antibody Dependent Cell Cytotoxicity (ADCC; macrophages and granulocytes). The artificial evolution of aptamers, to go into such adapters, against new viral variants will prevent a vaccine from ever becoming obsolete. Double-faced artificial polypeptides, like aptamers, have been computationally designed to re-direct humoral vaccine immunity from existing antibodies against other virus targets to SARS-CoV-2 by having one end of the polypeptide mimicking ACE2 viral receptor and the other a Hepatitis B antigen to bind the previous Hepatitis B vaccine induced antibody to the SARS-CoV-2 virus through the ACE2 viral receptor mimic (https://pubs.acs.org/doi/pdf/10.1021/acs.jpclett.0c03615).